
Abstract
Background: Glutathione is a key scavenging antioxidant that opposes the proinflammatory signaling of hydrogen peroxide. Boosting cellular glutathione levels may have broad utility in the prevention and treatment of disorders driven by oxidative stress. Supplemental N-acetylcysteine has been employed for this purpose. Could supplemental glycine likewise promote glutathione synthesis?
Methods: We conducted a review of the pertinent literature using PubMed.
Results: Tissue glycine levels are lower than the glutathione synthase Michaelis constant (Km) for glycine. When glycine availability is too low to sustain a normal rate of glutathione synthesis, the consequent rise in tissue levels of gamma-glutamylcysteine leads to an increase in urinary excretion of its alternative metabolite 5-L-oxoproline. The fact that urinary excretion of this metabolite is elevated in vegetarians and others consuming relatively low-protein diets strongly suggests that dietary glycine can be rate-limiting for glutathione synthesis in normally fed humans. Moreover, supplemental glycine has been reported to increase tissue glutathione levels in several animal studies. Glycine is a biosynthetic precursor for porphyrins, purines, creatine, sarcosine, and bile salts; is an agonist for glycine-gated chloride channels and a coagonist for N-methyl-D-aspartate receptors; inhibits protein glycation; and increases hepatic production of pyruvate, an effective scavenger of hydrogen peroxide. Supplemental glycine may have the potential for improving endothelial function, preventing cardiac hypertrophy, aiding control of metabolic syndrome, preventing the complications of diabetes, dampening inflammation, protecting the liver, and promoting effective sleep.
Conclusion: Clinical research is warranted to evaluate the impact of supplemental glycine on glutathione levels and on various health disorders.
- Acetylcysteine
- chloride channels
- glucagon
- glucagon-like peptide 1
- glutathione
- glutathione synthase
- glycine
INTRODUCTION
Reduced glutathione is one of the two most prominent scavenging antioxidants within cells alongside ascorbate; its concentration is in the low millimolar range. In addition to providing protection from nonspecific oxidant damage and acting as an electron donor for glutathione peroxidase and glutaredoxin, reduced glutathione reverses the prooxidative signaling mediated by hydrogen peroxide that plays a prominent pathogenic role in many health disorders.1-3 Glutathione is also employed in the conjugation and excretion of xenobiotics. Cellular levels of this key antioxidant tend to decline during the aging process.4 Therefore, practical strategies for optimizing tissue glutathione levels are needed.
METHODS
We conducted a review of the literature using PubMed keyword searches and citations in pertinent papers. Keywords were glutathione, glutathione synthase, N-acetylcysteine, glycine, chloride channels, 5-L-oxoproline, and lipoic acid.
DIETARY GLYCINE REGULATION OF GLUTATHIONE SYNTHESIS
Availability of the amino acid cysteine is known to be rate-limiting for the synthesis of glutathione, and it is well documented, both clinically and in animal studies, that cysteine supplementation—most practically achieved by administration of N-acetylcysteine (NAC)—can boost glutathione synthesis and levels.5,6 Dietary glycine levels may likewise modulate glutathione synthesis.
The tripeptide glutathione is synthesized in a 2-step process in which cysteine and glutamate are linked by glutamate cysteine ligase to form gamma-glutamylcysteine. This product is then linked to glycine by glutathione synthase. The Michaelis constant (Km) of human glutathione synthase for glycine has been reported as 0.58 mmol/L7 and 1.34 mmol/L8; why these values are not closer is not clear. A clinical study found that red blood cell concentrations of glycine averaged 487 μmol/L in young subjects and 218 μmol/L in elderly subjects; this level rose to 529 μmol/L in the elderly subjects after they were administered 100 mg/kg of both glycine and NAC daily for 14 days.9 Therefore, intracellular glycine concentrations appear to be no higher than, and possibly well below, the glutathione synthase Km for glycine, and practical glycine supplementation may notably increase intracellular glycine levels. Hence, glycine supplementation could possibly stimulate glutathione synthesis in humans. The expected rise in glutathione levels would likely be somewhat lower than the initial rise in synthesis rate, as glutathione acts as a feedback competitive inhibitor of glutamate cysteine ligase.10 In animal studies, glycine supplementation has been reported to increase tissue glutathione levels in milk-fed piglets and in rats subjected to burn injury, alcohol-induced liver injury, or fructose-induced metabolic syndrome.11-14
Clinical studies evaluating urinary excretion of 5-L-oxoproline provide evidence that dietary glycine is indeed limiting for glutathione synthesis in humans consuming their typical diets (as opposed to experimental diets depleted of glycine).15-17 When glycine availability is too low to sustain normal rates of glutathione synthesis, the consequent rise in tissue levels of gamma-glutamylcysteine results in increased conversion of this compound to 5-L-oxoproline that is then excreted in the urine.15,16 In a study comparing daily urinary excretion of this metabolite in omnivores and vegetarians, vegetarian males excreted 86% more 5-L-oxoproline than omnivore males, and vegetarian females excreted 37% more than omnivore females.17 These researchers also found that urinary excretion of 5-L-oxoproline rose significantly when subjects were switched from a diet providing 6.2 g of nitrogen per day to one providing 4 g of nitrogen per day. These findings strongly suggest that even though glycine is a nonessential amino acid that can be synthesized from other substrates, the amounts of glycine provided by some common diets is limiting for the synthesis of glutathione. Evidently, supplemental glycine should have its largest impact on glutathione levels in those who consume diets relatively low in protein, notably vegetarians and vegans.
These considerations suggest that intakes of cysteine and glycine jointly determine glutathione synthesis rates. Thus, at any given intake of cysteine, an increase in glycine intake could be expected to further boost glutathione synthesis and vice versa. Hence, joint supplementation with NAC and glycine could be expected to collaborate in stimulating glutathione production. In the previously cited clinical study,9 the elderly subjects received 100 mg/kg of both NAC and glycine for 14 days, and their red blood cell total glutathione levels (reduced glutathione plus diglutathione) rose from 1.26 mmol/L at baseline to 2.23 mmol/L, a concentration slightly higher than that measured in young subjects who did not take supplements.9 Moreover, the ratio of reduced glutathione to diglutathione rose from 7.4:1 to 16.1:1, indicative of a substantial improvement in redox status as confirmed by concurrent reductions in plasma markers of oxidative stress. The extent to which glycine contributed to the rise in cellular glutathione is unclear because the elderly subjects did not receive glycine or cysteine alone.
Tissue glutathione levels tend to decline with age, a phenomenon that may reflect declining function of Nrf2-mediated induction of phase 2 enzymes (including glutamate cysteine ligase), as well as lower tissue levels of cysteine and glycine.9,18-20 Phase 2 inducer nutraceuticals such as lipoic acid can be used to compensate for this decline in Nrf2 activity.18 An epidemiologic analysis found that while relatively low protein intakes predicted lower subsequent mortality in subjects <65 years, the opposite pattern was seen in those >65 years; greater mortality was seen in those in whom protein accounted for a lower percentage of total calories.21 Suboptimal cysteine intakes, exacerbating the age-related decline in glutathione levels, could play a mediating role in this phenomenon.4,22 An increase in glycine intake could potentially likewise have a beneficial impact on mortality rates in the elderly, most notably in those with low total protein intake.
HEALTH-PROTECTIVE POTENTIAL OF SUPPLEMENTAL GLYCINE
In addition to its role as a glutathione precursor, glycine is a substrate in the synthesis of porphyrins (heme), purines, creatine, sarcosine, and bile salts.23 Glycine constitutes approximately one-third of the amino acids in collagen and elastin and, hence, is a prominent component of connective tissues and the extracellular matrix. Additionally, glycine is an agonist for glycine-gated chloride channels that are expressed by a range of tissues peripherally and in the central nervous system.24-28 Activation of glycine-gated chloride channels has a depolarizing impact on cells that actively accumulate chloride and a hyperpolarizing impact on those that do not. Plasma glycine levels are close to the Km for glycine-gated chloride channels, so supplemental glycine, which can raise plasma glycine levels severalfold, tends to enhance the activity of these channels.29,30 Glycine is also a coagonist for certain neuronal N-methyl-D-aspartate (NMDA) channels.31,32 Additionally, glycine can suppress protein glycation and subsequent formation of advanced glycation end products by competing with protein-bound lysines for formation of Schiff bases with reactive aldehydes.33,34 In hepatocytes almost exclusively, glycine is metabolized to yield pyruvate (2 glycines generate 1 pyruvate); pyruvate can function as a scavenging antioxidant as it quenches hydrogen peroxide.35-39
GLYCINE AND METABOLIC SYNDROME
Owing to these diverse activities, supplemental glycine has shown intriguing and often beneficial effects in preclinical and clinical studies. Glycine provides protection from the adverse metabolic impacts of a high-fructose diet in rats, exerting favorable effects on insulin sensitivity, blood pressure, serum free fatty acids, and intraabdominal fat stores.40 Conceivably, this phenomenon reflects in part an upregulation of enteral production of glucagon-like peptide-1 (GLP-1).41 The L-cells that secrete GLP-1 express glycine-gated chloride channels; because L-cells accumulate chloride, activation of these channels exerts a depolarizing effect that promotes calcium influx and GLP-1 secretion.28 Although the impact of supplemental glycine on GLP-1 production has not been assessed in vivo, oral gelatin, notably rich in glycine, has been shown to boost GLP-1 levels.42,43 Glycine administered in a morning fasting metabolism has been shown to provoke increased glucagon secretion, likely reflecting a direct or indirect stimulatory effect on pancreatic alpha cells.44 Because both GLP-1 and glucagon boost hepatic fatty acid oxidation, these hormones might collaborate in mediating the protective impact of supplemental glycine on fructose-fed rats.41
Díaz-Flores et al administered 15-g glycine daily (5 g, 3 times daily) to patients with metabolic syndrome. Despite fasting glucose rising significantly from 101 mg/dL to 114 mg/dL (P=0.001), glycated hemoglobin fell from 7.81% to 6.45% (P=0.0001); the increase in fasting glucose in these patients might be explained by a stimulatory effect of glycine on glucagon secretion.45 Glycine supplementation was also associated with significant reductions in systolic blood pressure and plasma markers of oxidative stress.45 Importantly, when 5-g glycine was administered along with 25-g glucose in an oral glucose tolerance test, the subsequent increase in plasma glucose was notably blunted compared to the response to glucose alone. Because the glycine had relatively little impact on insulin secretion, the authors hypothesized that glycine was promoting intestinal production of an unknown hormone capable of boosting insulin sensitivity.44 When patients with type 2 diabetes were administered 5-g glycine 4 times daily for 6 months in an uncontrolled clinical trial, glycated hemoglobin fell from 9.6% to 6.9% (P<0.05); a favorable impact of glycine on oral glucose tolerance and on protein glycation might have collaborated in producing this benefit.46 A reduction of glycated hemoglobin in glycine-treated diabetic rats has also been reported.47 In rats with streptozotocin-induced diabetes, a favorable effect on certain diabetes complications (ie, glomerulosclerosis, cataracts, and microaneurysms of the retinal arteries) has been found.48-50
ANTIINFLAMMATORY EFFECTS OF GLYCINE
Macrophages, leukocytes, and Kupffer cells express glycine-gated chloride channels, and in these cells glycine exerts a hyperpolarizing effect, inhibiting calcium influx via voltage-sensitive calcium channels and thereby downregulating their proinflammatory activity.25,51,52 Studies have shown that dietary glycine can protect the livers of alcohol-fed rodents, and this effect is thought to reflect, in part, a suppression of Kupffer cell activation that lessens production of tumor necrosis factor alpha.53-56 Likewise, glycine administration improves liver status in rat models of nonalcoholic steatohepatitis induced by a high-fat, high-sugar diet or by methionine/choline deficiency.57-59 Increased hepatic production of pyruvate might contribute to the protection observed in these studies. Glycine supplementation is also beneficial in rodent models of inflammatory arthritis,60,61 although it had little impact on a canine model of osteoarthritis.62
GLYCINE AND VASCULAR HEALTH
Endothelial cells, and likely foam cells, express glycine-gated chloride channels.24 Glycine can exert a hyperpolarizing effect on endothelial cells that would enhance calcium influx.30,63 Endothelial cells lack voltage-sensitive calcium channels, so hyperpolarization therefore tends to drive calcium influx. Endothelial calcium influx activates nitric oxide synthase and also boosts expression of this enzyme via a mechanism dependent on a calcium/calmodulin-dependent kinase.64,65 Additionally, whereas membrane depolarization increases endothelial superoxide production via nicotinamide adenine dinucleotide phosphate (NADPH) oxidase activation, hyperpolarization has the opposite effect.66-68 Hence, supplemental glycine, via its hyperpolarizing effect on vascular endothelium and also via an antiinflammatory impact on foam cells, has been proposed to exert an antiatherosclerotic effect, a proposition that has not yet been tested.30 Consistent with this possibility, glycine administration to aging rats in drinking water for 2 months amplified endothelium-dependent vasodilation, increased mRNA expression of endothelial nitric oxide synthase, and downregulated expression of cyclooxygenase-2 and tumor necrosis factor alpha.69
Platelets likewise express glycine-responsive chloride channels; these channels have a hyperpolarizing action that suppresses calcium influx and promotes platelet stability.26
Ding et al measured plasma glycine levels in >4,000 patients who received coronary angiography for suspected angina pectoris.70 Higher levels of glycine were associated with a lesser extent of coronary atheroma and lower prevalence of obesity, hypertension, and diabetes. In light of the potential antiinflammatory effects of glycine, it is notable that C-reactive protein levels were nearly twice as high in patients in the bottom quartile of plasma glycine as in the upper quartile (2.48 vs 1.35 mg/L, P<0.001). During a median follow-up of 7.4 years, 616 patients experienced myocardial infarcts. After multivariate adjustments for known risk factors, patients whose glycine was in the fifth quintile at baseline were 29% less likely to have experienced an infarct than patients in the first quintile (hazard ratio 0.71; 95% confidence interval 0.54-0.94, P=0.016) (Figure and Table). Whether these associations reflect cause-effect relationships remains to be determined, but the findings are consistent with the possibility that elevated glycine levels provide protection from cardiovascular events. Of related interest is a report that dietary glycine prevents development of cardiac hypertrophy in rats subjected to pressure overload.71 This effect may be mediated by glycine receptors expressed on cardiomyocytes.27,71
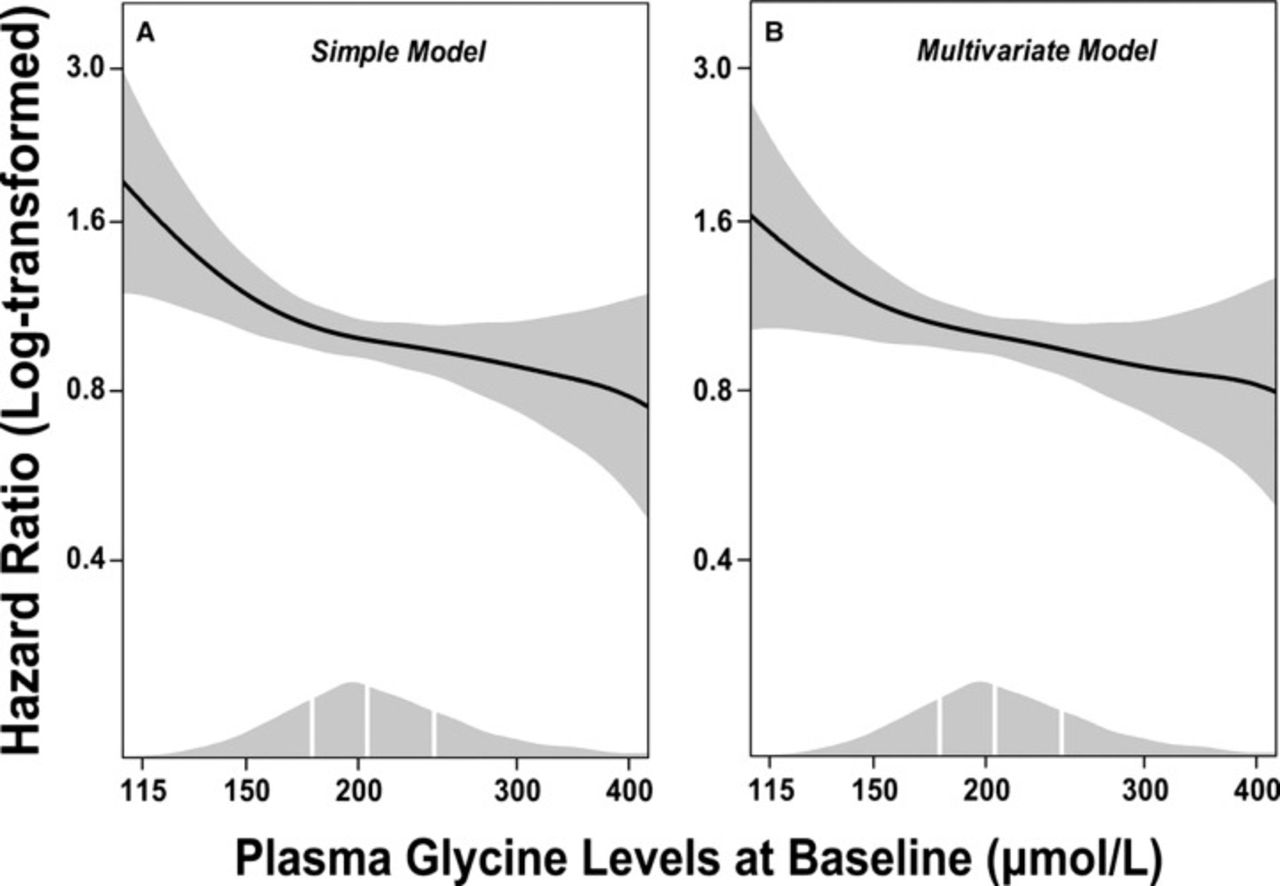
Dose-response associations between (log-transformed) plasma glycine and risk of acute myocardial infarction. Generalized additive regressions are used with the adjustment for age and sex in the simple model (A) and with additional adjustment for smoking, obesity, hypertension, diabetes mellitus, angiographic extent of coronary artery disease (ordinal), estimated glomerular filtration rate, apolipoprotein A-1, apolipoprotein B, and statin treatment in the multivariate model (B). The solid lines and the shaded areas represent hazard ratios of plasma glycine and their 95% confidence intervals, respectively. The areas under the curve along the x-axes represent the distributions of the plasma glycine concentrations (μmol/L) in the total population. The vertical white lines denote the 25th, 50th, and 75th percentiles of plasma glycine, respectively. (Republished with permission according to a Creative Commons Attribution-NonCommercial license; Ding et al.70)
Hazard Ratios of Acute Myocardial Infarction According to Quintiles of Plasma Glycine Levels
IMPACTS ON CANCER, CONNECTIVE TISSUE, AND THE CENTRAL NERVOUS SYSTEM
Glycine might prove to have some utility in cancer management. Indeed, glycine supplementation has been reported to slow the growth of various implanted tumors in rodents, likely owing to an antiangiogenic impact on endothelial cells.72-74 However, this effect might modestly retard wound healing.73 The antiangiogenic effect of glycine may reflect the fact that activation of endothelial NADPH oxidase, which the hyperpolarizing action of glycine may oppose, is a key mediator of the signaling pathway whereby vascular endothelial growth factor and its Flk-1 receptor promote proliferation and migration of endothelial cells.75,76 Additionally, glycine acts directly on a human hepatocyte line, via glycine receptors, to inhibit expression of vascular endothelial growth factor; proliferation of these cells was not influenced by glycine.77 The antiinflammatory effects of glycine may extend to cancer cachexia, as glycine administration has been found to blunt the loss of muscle and fat mass and decrease inflammatory markers in mice implanted with C26 cancer cells.78
The possibility that increased glycine intake may have a favorable impact on bones and tendons is being evaluated. In vitro, glycine was found to stimulate the proliferation of an osteoblast cell line; when administered to ovariectomized rats, glycine favorably influenced bone mineral density and ultrastructure.79 In a clinical study evaluating monozygotic twins, the twins with a higher dietary intake of glycine or alanine tended to have higher spinal bone mineral density.80 In a rat model of Achilles tendinitis, supplemental glycine aided healing.81-83 The fact that collagen and elastin are remarkably rich in glycine suggests that optimal glycine nutrition might be beneficial for the health of connective tissues (including the cardiovascular system).23
Finally, glycine, via its interaction with glycine and NMDA receptors, has potential to influence the central nervous system.
A controlled clinical trial found that glycine supplementation prior to bedtime improves sleep quality in patients afflicted with insomnia.84 However, daytime administration of glycine did not cause drowsiness.85 In rats, glycine administration boosts non–rapid eye movement sleep and amplifies the decline in body temperature (reflecting cutaneous vasodilation) that typically accompanies sleep. These effects were found to be mediated by NMDA receptors in the suprachiasmatic nucleus.86 Glycine has also been evaluated as an adjuvant to neuroleptic treatment of schizophrenia; both high-dose glycine and D-serine, a full agonist for the glycine-binding site on NMDA receptors, have been reported to aid control of negative symptoms.87,88
GLYCINE AS A SUPPLEMENTAL NUTRIENT
Clinical studies of supplemental glycine that have achieved interesting results have used daily glycine doses ranging from 3 g to as high as 0.8 g/kg, split into 2-4 servings.45,46,85,89 These studies have failed to note any significant side effects, presumably reflecting the fact that glycine is efficiently absorbed and hence does not cause diarrhea.45,46,85,89 Ample doses of glycine are not impractical because glycine is inexpensive, has a mildly sweet flavor, is highly and rapidly soluble, and can be added to a wide range of beverages.
CONCLUSION
We have good reason to suspect that clinically feasible doses of oral glycine can boost tissue levels of glutathione, an effect that could be amplified by concurrent administration of NAC and/or a phase 2 inducer such as lipoic acid. Because glutathione levels decline during the aging process, supplemental glycine would likely have its most notable impact in elderly people with diets relatively low in protein. Moreover, by serving as a precursor for a range of biomolecules, by acting as an agonist for glycine-gated chloride channels and a coagonist for NMDA receptors, and by suppressing protein glycation, increased intakes of glycine have the potential to exert a broad range of protective effects: inhibiting atherosclerosis and cardiac hypertrophy, controlling metabolic syndrome, aiding control of diabetes and its complications, protecting the liver exposed to excess alcohol or fatty acids, blunting inflammation, and aiding sleep quality. Clinical research is warranted to evaluate the impact of supplemental glycine on glutathione levels and on various health disorders.
This article meets the Accreditation Council for Graduate Medical Education and the American Board of Medical Specialties Maintenance of Certification competencies for Patient Care and Medical Knowledge.
ACKNOWLEDGMENTS
Mark F. McCarty is coowner and science director of a nutraceutical company that sells a product containing glycine. James H. O'Keefe is the chief medical officer and founder of CardioTabs, a nutraceutical company. James J. DiNicolantonio has no conflicts of interest to disclose.
- © Academic Division of Ochsner Clinic Foundation 2018
REFERENCES
In this issue








{kind=link}
Jump to section
- Article
- Abstract
- INTRODUCTION
- METHODS
- DIETARY GLYCINE REGULATION OF GLUTATHIONE SYNTHESIS
- HEALTH-PROTECTIVE POTENTIAL OF SUPPLEMENTAL GLYCINE
- GLYCINE AND METABOLIC SYNDROME
- ANTIINFLAMMATORY EFFECTS OF GLYCINE
- GLYCINE AND VASCULAR HEALTH
- IMPACTS ON CANCER, CONNECTIVE TISSUE, AND THE CENTRAL NERVOUS SYSTEM
- GLYCINE AS A SUPPLEMENTAL NUTRIENT
- CONCLUSION
- ACKNOWLEDGMENTS
- REFERENCES
- Figures & Data
- References
- Info & Metrics